
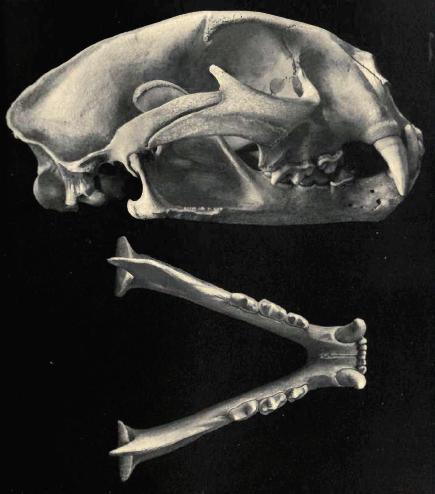
Skull of Puma concolor, image from here
A cheetah stands yawning. Its shoulders shiver as it makes a feeble attempt to get rid of some irritating flies. Nearby, two spotted cubs hide under a bush to keep them safe from roaming lions, while a herd of antelope graze. Can you picture it? Would it surprise you to learn that I am describing a speculative scenario set in the late Pleistocene of Wyoming? The lithe predator species here is Miracinonyx trumani, the American cheetah, not the African cheetah (Acinonyx jubatus). The lions menacing those cubs are American lions (Panthera leo atrox) not African lions (Panthera leo leo). And the antelopes are the supremely weird pronghorns (Antilocapra americana).
The American cheetah is perhaps the most enigmatic of felid taxa. No one had any idea it existed until the 1970s! Prior to this, material we now know belongs to M. trumani was assumed to be a kind of puma or was so fragmentary it was mistaken for other Pleistocene Felidae.
This all changed when the pitfall site of Natural Trap Cave was excavated [currently being re-excavated- follow #NTCave14 on Twitter]. At the bottom of this inescapable deathtrap, excavators found the remains of mammoth, lions, bighorn sheep, camels, and bison, as well as thousands of smaller animals. Crucially, as the cave is over 80 feet deep (measured from the surface) any creature that falls in is instantly killed (or severely incapacitated until death), with no escape and no potential for post-mortem disturbance. This has led to many articulated and associated remains being found, the most important of which were from Miracinonyx. From these remains, researchers were able to show that the animal had a whole suite of features that separated it from puma. These included spectacularly long legs, a shortened skull with enlarged nares, and specialised features of the fibula. Initially, researchers assumed that these adaptations must be due to a shared ancestry and placed Miracinonyx as the sister species to Acinonyx, implying that the African cheetah may have been just another New World endemic that migrated back to the old world (like dogs, horses, and camels). Later research tied all these specialisations into convergently evolved adaptations to a highly cursorial lifestyle. In effect, the American cheetah looked like the African cheetah because they had both evolved to be extremely fast, pursuit predators, and not due to a close phylogenetic relationship. The final piece in the puzzle of Miracinonyx origin came in 2005, when I was lucky enough to extract some ancient DNA from a piece of M.trumani humerus from Natural Trap Cave, dated at 19,765±80 radiocarbon years old. With a lot of help from colleagues and friends, we did some phylogenetic analyses that showed with a high degree of statistical support that Miracinonyx trumani and Puma concolor were sister species, that diverged approximately three million years ago. Our initial conclusion was that perhaps the divergence date was significant, as this was approximately the time when grassland prairie started to expand in North America, providing opportunities for a savannah predator to evolve.
However, there may be a bit of a problem with this straightforward scenario. Bones discovered within caves at the Grand Canyon have recently been attributed to Miracinonyx. Could the “American cheetah” have been more like an “American snow leopard”, hunting on the scree and sheer cliffs of this mountainous environment? I am not completely convinced, although the suggestion is very intriguing. The Grand Canyon material was only compared to modern puma and Pleistocene Miracinonyx material and grouped with Miracinonyx, mostly on the basis of larger size. However, we know that Pleistocene puma were much larger than modern day puma, and if the canyon material could be compared to a dataset that included Pleistocene puma I am sure they might provide a better match. The puma is perhaps the most versatile large cat in the world and can hunt in habitats as diverse as tropical rainforest and Canadian tundra. Canyon living would be no problem.

The jaguarundi (or eyra). This housecat sized South American endemic is sometimes tamed as a pet by local Quechua groups. Image from here.
So, if Miracinonyx trumani and Puma concolor evolved from a common ancestor, where did that cat come from? The puma lineage consists of three living cats (as well as numerous extinct ones). The most basal member is the cheetah, found in Africa and Asia (there is still a small population of cheetah in Iran), while the puma and the jaguarundi (Herpailurus [Puma] yagouaroundi) are found in the New World. There is in fact a potential ancestor for the Miracinonyx, puma and jaguarundi known from the Old World: Puma pardoides. This strange cat has been found in various sites all over Eurasia and could possibly be the starting point for jaguarundi, puma, and Miracinonyx.
A small note on the etymology of Miracinonyx. It comes from a compound of the Latin “mirus” (=astounding, surprising), and the linnean genus for cheetah “Acinonyx” (which itself comes from the Greek prefix a- and -kineo, to move: a reference to the supposedly non-retractile claws in cheetah). The American cheetah line died out at the same time as American lions, sabretooths, and ground sloths at the end of the Pleistocene.
Written by Ross Barnett (@DeepFriedDNA)
Further Reading:
Miracinonyx trumani from the Grand Canyon. Jean-Paul Hodnett
Did false cheetahs give pronghorns a need for speed? Brian Switek (@laelaps)
Big spotted pumas… Miracinonyx redux?Darren Naish (@TetZoo)
Adams, D. B. “The Cheetah: Native American.” Science 205 (14 September 1979): 1155-58. [Article]
Barnett, R., I. Barnes, M. J. Phillips, L. D. Martin, C. R. Harington, J. A. Leonard, and A. Cooper. “Evolution of the Extinct Sabretooths and American Cheetahlike Cat.” Current Biology 15, no. 15 (2005): R589-R90. [Article]
Hemmer, H., R. D. Kahlke, and A. K. Vekua. “The Old World Puma – Puma Pardoides (Owen, 1846) (Carnivora : Felidae) – in the Lower Villafranchian (Upper Pliocene) of Kvabebi (East Georgia, Transcaucasia) and Its Evolutionary and Biogeographical Significance.” Neues Jahrbuch für Geologie und Paläontologie 233, no. 2 (Aug 2004): 197-231. [Article]
Kurtén, B. “Fossil Puma (Mammalia:Felidae) in North America.” Netherlands Journal of Zoology 26 (1976): 502-34. [Article]
Martin, L. D., and B. M. Gilbert. “Excavations at Natural Trap Cave.” Transactions of the Nebraska Academt of Sciences and Affiliated Societies VI (1978): 106-16. [Article]
Martin, L. D., B. M. Gilbert, and D. B. Adams. “A Cheetah-Like Cat in the North American Pleistocene.” Science 195 (1977): 981-82. [Article]
Morgan, G. S., and K. L. Seymour. “Fossil History of the Panther (Puma Concolor) and the Cheetah-Like Cat (Miracinonyx Inexpectatus) in Florida.” Bulletin of the Florida Museum of Natural History 40, no. 2 (1997): 177-219. [Article]
van Valkenburgh, B., F. Grady, and B. Kurtén. “The Plio-Pleistocene Cheetah-Like Cat Miracinonyx Inexpectatus of North America.” Journal of Vertebrate Palaeontology 10, no. 4 (December 1990): 434-54. [Article]




























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.JPG){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
#mediaviewer/File:Pelotes_r%C3%A9jection_Asio_Otus.jpg){kind=link}
{kind=link}