
Ask anyone to think of Pleistocene beasts and they will invariably conjure up images of herds of woolly mammoth, bison and horses sweeping majestically across the grassy plains, perhaps with a pack of dire wolves or lone short-faced bear stalking them at a distance. Indeed, it is these charismatic animals, with their large and impressive skeletons now reconstituted in museums across the world, that have inspired many young children to go on and become paleontologists later in life.
But as that person continues to dream of the Pleistocene, little thought will likely be spared for the beasts within, such as Strongyloides shastensis and Archeococcidia nothrotheriopsae. Their diminutive size does not lend well to impressive museum displays or awe-inspiring dioramas, and there is slim chance that they are on anyone’s re-wilding wish lists. And, as someone who has spent the past decade studying extinct megafauna, I have been as guilty as anyone in overlooking them in the past. But now they deserve a voice, for it is beasts such as these that could actually represent the majority of biodiversity lost in global late Quaternary extinction events.
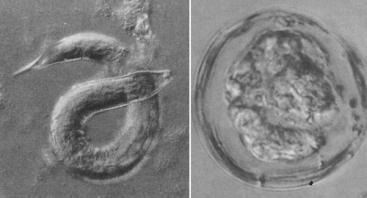
The nematode Strongyloides shastensis and coccidian Archeococcidia nothrotheriopsae once foraged in the digestive tracts of Shasta ground sloth. Their habitat disappeared at the end of the Pleistocene, and as a result they is now likely extinct. Images from Schmidt & Duszynski (1992).
Strongyloides shastensis and Archeococcidia nothrotheriopsae were both gut parasites of the Shasta ground sloth, first described by Gerald Schmidt and Donald Duszynski in 1992. Parasites are simply a fact of life for most wild animals. From an invertebrate biologist’s viewpoint, the charismatic megafaunal beasts of the Pleistocene were simply mobile habitats, providing shelter and food for diverse communities of smaller organisms. These included a terrifyingly large array of different beasts including nematodes (roundworms), trematodes (fluke worms), coccidians and hepatazoons (both single celled parasites), lice, and mites. Some of these organisms would have lived on external surfaces of their hosts (e.g. lice) while others would have been found in various organs, the bloodstream, or gastrointestinal tract. Many of these organisms were likely host-specific, that is, they could only have survived on, or in, a single host species. But having lives that were so intertwined with particular host-species also created obvious issues. For example, what would happen if those species were to suddenly go extinct?
Loss of mutualistic, or parasitic species, along with their partners is a process known as ‘co-extinction’ and it has been suggested that this process may be the main driver of biodiversity loss during mass extinction events such as those of the late Quaternary (see Koh et al. 2004, Dunn 2009, Dunn et al. 2009). However, studying late Quaternary co-extinctions clearly has its challenges. Small organisms without hard body parts are under-represented in the fossil record, and the dependence of parasites on their hosts means they are seldom found in the external environment. As a result, little is known about exactly how prehistoric parasites responded to habitat loss. Well-documented cases of even relatively recent parasite co-extinctions (e.g. the huia louse, Rallicola extinctus) are rare, due to the challenges of taxonomic identification of parasites and the difficulty of proving parasite host-specificity.

The extinct New Zealand huia (Heteralocha acutirostris) and the equally extinct louse (Rallicola extinctus) that once lived on the huia’s feathers. (Image from here)
Coprolites (ancient, fossilised dung) may provide a key to understanding the co-extinction process, by offering a unique window into the lives and deaths of some parasites; such as those that inhabited the gastrointestinal tracts of megafauna. In fact, it was coprolites of the Shasta ground sloth, preserved for millennia in Arizona’s Rampart Cave, that allowed Schmidt and Duszynski the first insights into the micro-beasts that were once associated with that species. By digging in the poo, they found the tiny parasites.
More recently, myself and colleagues have begun to investigate parasites in coprolites from the moa, the large extinct ratite birds of New Zealand. In our first study published last year (Wood et al. 2013) we were able to show that with a reasonable sample size of coprolites assigned to different moa species using ancient DNA identification, it is possible to determine with some degree of confidence whether the parasite taxa seen in the coprolites were host-specific or not (and several were).
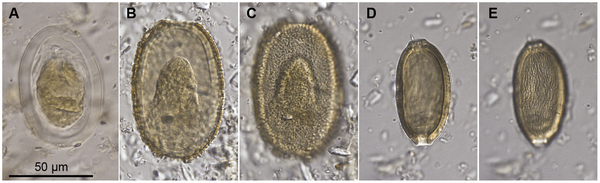
Eggs of parasitic nematodes isolated from coprolites of the giant New Zealand moas. (Images from Wood et al., 2013)
With recent advances in techniques for extracting ancient DNA from coprolites, and rapidly developing DNA sequencing technologies, we likely now have the opportunity to examine Pleistocene co-extinctions in greater detail than has previously been possible. Such research may help broaden our understanding of late Quaternary extinction events, allowing us to look not only at extinctions in vertebrate faunas, but also into the cryptic, and potentially more dramatic, consequences for invertebrate communities. So, the next time you picture a mammoth wandering through the ice and snow, spare a thought for those who went along for the ride!
Written by Jamie Wood (@larusnz)
Edited by Jan Freedman (@janfreedman)
Further Reading:
Bush, A.O., Kennedy, C.R. (1994) ‘Host fragmentation and helminth parasites: Hedging your bets against extinction.’ International Journal of Parasitology 24, 1333–1343. [Abstract only]
Dunn, R.R. (2009) ‘Coextinction: anecdotes, models, and speculation’. In: Turvey S, editor. Holocene extinctions. Oxford: Oxford University Press. 167–180. [Book]
Dunn, R.R., Harris, N.C., Colwell, R.K., Koh, L.P., Sodhi, N.S. (2009) ‘The sixth mass coextinction: are most endangered species parasites and mutualists?’ Proceedings of the Royal Society of London Ser B 276, 3037–3045. [Full article]
Ferreira, L.F., Araujo, A., Duarte, A.N. (1993) ‘Nematode larvae in fossilized animal coprolites from Lower and Middle Pleistocene sites, Central Italy’. Journal of Parasitology 79, 440–442. [Full article]
Koh, L.P., Dunn, R.R., Sodhi, N.S., Colwell, R.K., Proctor, H.C., et al. (2004) ‘Species coextinctions and the biodiversity crisis.’ Science 305, 1632–1634. [Abstract only]
Schmidt, G.D., Duszynski, D.W. (1992), ‘Parasites of the extinct Shasta ground sloth, Nothrotheriops shastensis, in Rampart Cave, Arizona’ Journal of Parasitology 78. 811–816. [Full article]
Wood, J.R., Wilmshurst, J.M, Rawlence N.J., Bonner, K.I., Worthy, T.H, Kinsella, J.M., Cooper, A. (2013), ‘A Megafauna’s Microfauna: Gastrointestinal Parasites of New Zealand’s Extinct Moa (Aves: Dinornithiformes)’ PloS ONE 8(2). E57315. [Full article]


{kind=link}
Pingback: 2015: A review of the Beasts | TwilightBeasts
Pingback: The most (and least) read posts of 2022 | TwilightBeasts