
Our planet’s climate fluctuations move to the slow, almost imperceptible dance steps of tilts in our axis and wobbles in our precession around the star we call the Sun. These movements which cause natural global-scale climate changes are known as the Milankovitch Cycles, and they take a long time in human terms – a brief warm interstadial followed by a big freeze every 100,000 years.
Before the Pleistocene, there were many epochs of warm, and cold, wet and dry. Some were cataclysmic to the animal and plant life of that period, creating mass extinctions and subsequently allowing niche species to replace them. So, before the first rays of icy sunlight shone upon the Pleistocene creatures usually hosted on Twilight Beasts, there were other animals, equally fascinating, which existed in the even stranger world of the Pliocene, which dates between 5.3 to 2.6 million years ago. It was a time of seasonally fluctuating polar ice caps until the icy wind of the Pleistocene issued in the ice age proper; it was a time when the Mediterranean Sea dried out to become a grassland, only to be flooded again and become the sea we know today – this is known as the Zanclean Flood. Although temperatures were slightly warmer than the present day, the Pliocene’s cooler, drier climate contributed to the globally proliferating grasslands. It was a time of beginnings, endings and changes…. and horned gophers.
Gentle reader, you read this correctly; A gopher, but no ordinary one – the Austin Powers, rodent of mystery. This rodent was, literally, horny ( Yeah baby – oh, behave!).
No kidding – it actually had a pair of little horns.

Mummy Ceratogaulus with wee ones (Image from PalaeoPastori)
Let me introduce you to one of the real oddities of our extinct bestiary – Ceratogaulus. A gopher-like beastie, adorable as all little scurrying things are, but with slightly silly looking little devil horns on their noses, and quite a mysterious little beast in its own right.
Technically, they were closer to ground squirrels and marmots, being Sciuromorphs, of the Mylagaulidae clade, but as we know, things aren’t always named what they actually are (cough… cough… looking at Megaloceros and that pesky elk labelling!). They were not particularly large creatures, on average around 30cm in length, though some appear to have been larger, and they once scurried and scraped the Badlands of Nebraska in North America. There were four definite species of Ceratogaulus – C. rhinosauros, C. hatcheri, C. minor and C. anecdotus and a newly discovered one in 2013, which likely is the fifth, C. robustus, which has only been found at Skull Ridge, New Mexico so far. Sadly for us and the internet (for this rodent would be a star on Twitter, Instagram et al if it was around today) the entire clade are extinct. They were the smallest known mammals to ever have horns, and nobody is really quite sure why.
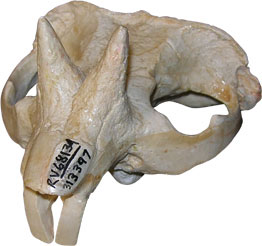
The thick horns can be seen clearly on this skull. (Image from http://www.ucmp.berkeley.edu/museum)
Way back in the early years of the 20th century, when the discovery of our ratty pal was still fresh in everyone’s minds from the 1890’s, it was taken as accepted that those horns were needed for burrowing – dig that horn in, shake that stocky wee neck around, and – scoop! The earth moves! There was even a proposed theory that the fossilised Daimonelix tunnels found during excavations across Nebraska (sometimes known as ‘Devils Corkscrews’) were the remains of their underground lairs. Well, those twisting and convoluted burrows turned out to be the homes of totally different critters of the Miocene/Pliocene – Palaeocastor fossor, which was a giant ancient beaver – but that’s a story for a different day!
It all would have been a fine and likely theory if the horns were not positioned where they were (right on the nose) and pointing upwards. Any pressure applied from moving earth, or tunnelling would have broken those substantial horns. No, those well-developed rodent claws would have made quite good burrows with or without any protuberances on the schnozz.
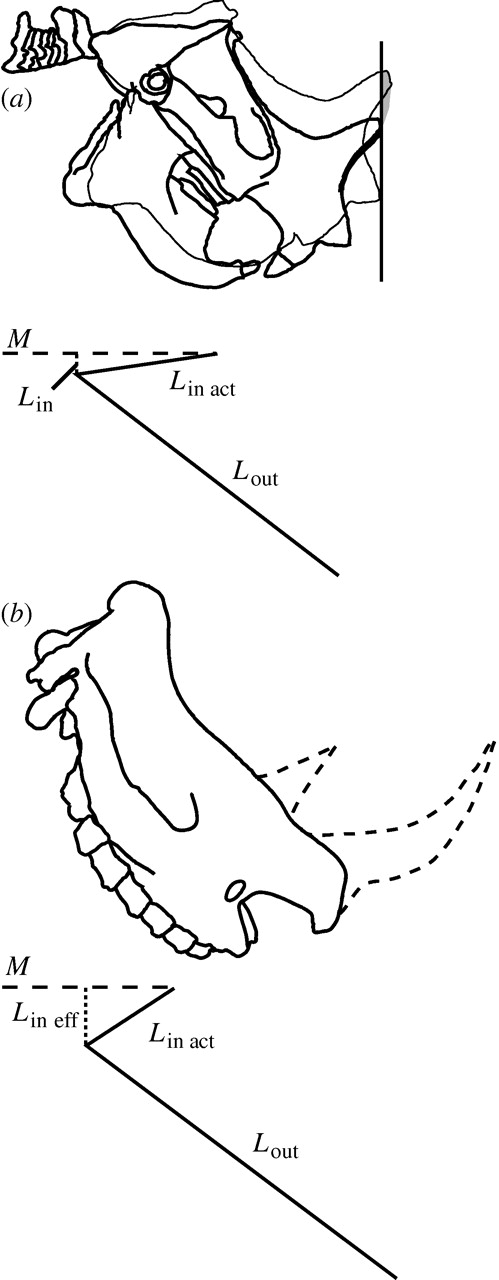
Reasons why Ceratogaulus could not have been very successful at shifting earth with those horns – ouch! ( from Hopkins 2005)
Could those horns have been indicators of sexual dimorphism? Nope, the lady Ceratogaulus had them as well, so that theory doesn’t work. Could the horns have been some sort of sexual display, the way deer use their antlers? The fight for love between rival male Megaloceros would have been a phenomenal thing to see. Imagine the ermm, ‘terrifying’ crash of horns as passion-crazed horny gophers of the Pliocene slugged it out for damsels fair. Well, it’s possible, but would be a bit of an outlier for similar reasons to the digging hypothesis.
The answer most consider to be right in the present day (unless a new idea is put forward) is that basically, Ceratogaulus was too large to be underground all the time, and would have spent a fair bit of time scratching around for food. Rodents all too often are nature’s easy snack-pack for carnivores, and being roughly marmot-sized, with short legs and a chubby round body, our horny rodent wouldn’t have been able to run anywhere very fast, unlike a tiny wee mouse or a tree dwelling squirrel. Hence, perhaps those horns. A well-aimed head-butt with those formidable bony horns and a peckish predator would be pretty sore, and possibly deterred from pursuing a would-be lunch of rodent.
And they were a successful species, extending across the Miocene into the Pliocene, but there were changes afoot, and none of it, at this time, anything whatsoever to do with humans messing up. Climate changes, and attendant vegetation cover, were more likely the cause of Ceratogaulus not making it into the era of the Twilight Beasts, or beyond.

Small but deadly, our little Ceratogaulus would have been a tough prey for many predators. (image Jan Freedman)
During the Miocene period, the prairies of North America gradually became more open and grassy, as a response to the cooling temperatures and decreased precipitation, and by the Pliocene period the balance tilted in favour of grazing animals rather than browsers, as woodland suffered in many areas. As bad luck would have it, one of the areas which succumbed to the loss of deciduous woodland was Nebraska and the Great Plains region of North America. Small creatures had increasingly less places to hide.
The Pliocene, as I said at the start, was a time of beginnings, endings and change – even the grasslands which now stretched across America photosynthesised differently, as C3 and C4 grasses react in very different ways to sunlight and carry different contents of nutrition as a result. As populations of small creatures like Ceratogaulus declined, so too did biodiversity, as these little ones are seed dispersal agents for nature, carrying seeds in their fur, or popping them out in their poo. Nature is balanced carefully with such debits and credits. When one factor goes wrong in an ecosystem, everything slips. The earth was gradually dipping into a cooler phase, making way for the lumbering glaciers of the Pleistocene.
As the changes of the late Pliocene were happening on the North American continent, in drought-parched Africa a little hominin was learning to stand up, use simple tools, and almost certainly wonder what lay beyond the parched horizon. The rise and evolution of that small creature would change the face of the Earth in the coming millennia arguably even more than the ice sheets gathering in the polar regions by the end of the Pliocene. Its children’s’ children and beyond would be responsible for exterminating many other creatures on the planet. But that was the future….
So, there you have it , a creature so fantastical it likely belongs more in an X-File, alongside jackalopes, than in a palaeo bestiary; a ground squirrel fluff-ball with little horns. No memory of these creatures passed into mythology or folklore, for no human ever witnessed their rotund little bodies mooching through for juicy herbs, roots, berries and bugs, or whatever else they loved to eat. Probably a good thing, too – sadly we know what humans are capable of. No cave art records their sunrise, or their sunset. We have just been left with wonder, wishing we could have petted those little heads, and maybe more than anything else – find out what those horns were actually for!
Written by Rena Maguire (@JustRena)
Further Reading:
Emlen, D. J. (2008). ‘The evolution of animal weapons’. Annual Review of Ecology, Evolution, and Systematics. pp.387-413. [Full article]
Hopkins, S. S. (2008). ‘Reassessing the mass of exceptionally large rodents using toothrow length and area as proxies for body mass’. Journal of Mammalogy, 89 (1), pp. 232-243. [Full text]
Hopkins, S. S. (2005). ‘The evolution of fossoriality and the adaptive role of horns in the Mylagaulidae (Mammalia: Rodentia)’. Proceedings of the Royal Society of London B: Biological Sciences, 272 (1573), pp. 1705-1713. [Full article]
Korth, W. W. (2013). ‘Mylagaulid Rodents (Mammalia: Rodentia: Mylagaulidae) from the Middle Miocene (Barstovian) of New Mexico’. Annals of Carnegie Museum. 81 (4). pp. 233-245. [Full article]
Meyer, R. (1999). ‘Helical burrows as a palaeoclimate response: Daimonelix by Palaeocastor’. Palaeogeography, Palaeoclimatology, Palaeoecology. 147. pp 291–298. [Full article]
Schmerge, J. D. (2015). ‘Interpretation of Euhapsine (Castoridae: Palaeocastorinae) Burrowing Behaviors Based on the Functional Anatomy of the Teeth and Skull with a Description of a New Species and Genus’. [Full article]
Strömberg, C &. McInerney, F. (2011). ‘The Neogene transition from C3 to C4 grasslands in North America: assemblage analysis of fossil phytoliths’, Paleobiology. 37 (1). pp 50-71. [Full article]
Stromberg, C. (2011). ‘Evolution of grasses and grassland systems’. Annual Review Earth and Planet Sciences. 39. pp. 517-544. [Abstract only]
Webb, S. D. (1966). ‘A relict species of the burrowing rodent, Mylagaulus, from the Pliocene of Florida’. Journal of Mammalogy, 47 (3) .pp. 401-412. [Abstract only]


Reblogged this on Shallow Thinking.
Pingback: Small but mighty – clearancerose
Fascinating. A side thought – reminded me of the dangerous rabbit in Monty Python and the Holy Grail.
Neat! 🙂
Pingback: The Rise and Reign of the Mammals | TwilightBeasts