
Nothing beats the thrill of holding a fossil in the palm of my hand. Feeling the smooth cold surface gently resting on the top of my bare skin instantly transports me back to the time that creature was alive: for I am holding the remains of something that breathed, moved and ate. A reindeer hoof, with its slightly strange curved triangular shape, takes me back to hearing the soft crunch of the frozen snow as the padded feet walk across the desolate land. Holding a Woolly Rhinoceros tooth, I can see the huge beast snorting blasts of frozen air through its nose as the big lips pull up chunks of grass and the creature munches loudly.
Each fossil tells its own story. That fossil found in that place provides us with a rich amount of information. Woolly Rhinoceros and Woolly Mammoths tell of a time when it was cold, very cold – not too different from the Siberian tundra today. Hippopotamus fossils take us even further back in time to when temperatures were much hotter than they are today. These megafauna along with sediment they were preserved in, plants, and pollen all recreate a world long vanished. And in surprising detail too. From careful excavations around the world we have a fairly good picture of the Pleistocene and how local environments have been changing in the past. And what’s even more exiting is we are still learning more and more every year.

A gorgeous sub-fossil hyena jaw from Kitley Caves, Devon from the collections at Plymouth City Museum and Art Gallery.
The Pleistocene is often referred to as the ‘Ice Age’ giving the misconception of a long period of freezing temperatures, and enormous glaciers reaching down on Europe and North America. From the large fossils we know that between 2.6 million years ago until around 11,700 years ago, there were times when the planet was colder, and sometimes times it was warmer. Those cold times (the glacial periods), allowed Mammoths to lollop across Devon. Warmer spells (interglacial periods) were warm enough for hippopotamus to be very comfortable living in Yorkshire! The Pleistocene, then, was a time when the climate fluctuated between extreme warm and rather cold periods.
This simple picture of a couple of pulsating glaciers punctuated by tropical paradises is far, far, far from reality. To find out just what the climate of the Pleistocene was like we look not to the land, but the oceans. And within it, silently drifting along the undercurrents is a tiny beast that holds the keys to unlocking the secrets of the past.
Foraminifera are teeny, tiny singled celled organisms that grow a shell (called a test). They are mainly marine organisms, with a few species floating around in freshwater environments. Many live in the muddy sediment, while others float in the water nearer the surface, with a pretty cool feeding strategy: the walls of the cell are loose, and it is this ectoplasm that swings freely like miniature arms to catch tiny floating particles which it absorbs as food. Pretty nifty little things. And most of the 4000 or so species are small, as small as this full stop. (One species is a huge single celled beast that would fit in my outstretched hand.) What make foraminifera special is their shells. And it is these shells that hold the secrets to the past.

The tests of four different species of forams imaged under a Scanning Electron Microscope highlighting incredible detail. (Image Public Domain)
A little recap on some very simple chemistry reveals why the shells of our smallest Twilight Beast are so important. A water molecule contains two atoms of hydrogen bound with one atom of oxygen. An atom can have a different number of neutrons and still be the same atom: all that changes is the density of that atom. So if an Oxygen atom had just 8 neutrons and 8 protons (16O)this atom would be lighter than an Oxygen atom with 10 neutrons and 8 protons (18O). Water molecules in the ocean are different from each other with some being ‘lighter’ and others being ‘heavier’. (The number of neutrons makes the Oxygen Isotope lighter or heavier: the number of protons remains the same). And this is where our miniature beasts come in.
As the foraminifera grow their shells, they absorb oxygen out of the water, which is trapped in their shells. The forams take the ratio of light Oxygen isotope (16O) to the heavier Oxygen isotope (18O) to their graves – literally. When these little organisms die, they float to the bottom of the ocean, preserving in their thin little shells the ratio of the Oxygen isotopes from the oceans when they were alive. This ratio of the light Oxygen isotope (16O) to the heavier Oxygen isotope (18O)is important as it can tell us about the temperature of the planet. And here’s how…
We know the heat from the sun evaporates water. What is really cool is that the water molecules with the smaller number of neutrons (16O) are evaporated first, because they are lighter. Normally, the water returns to the oceans through rivers, so the Oxygen isotope ratio of 16O and 18O remains equal. But, during colder glacial periods more of the water falls as snow, and gets trapped in the growing glaciers so the oceans have more 18O than 16O. And the amazing thing is that this is recorded in the microscopic shells of foraminifera. What’s more, it can be measured.
Taking core samples from the ocean sediment is opening up a window into the past. And through this window we are seeing a past that rapidly fluctuated between long periods of cooling (glacial) terminating in fast shorter periods of extremely hot climates (interglacials). At the top of the sediment core is today, the present, and as you run your finger down the muddy core, you are tracing your finger back in time. The core is sampled for forams at regular spots, and the Oxygen isotopes in the tests are tested, providing a zig-zag style graph where the ratio has changed through the core sample. But something is missing. Time. Sure, we know that as you go deeper in the sediment core you are going furrther back in time. But how far back? Weeks? Years? Millennia?
The sediment core sample can be dated by radiocarbon dating, uranium-thorium dating, or correlating to already dated events (like magnetic reversals and volcanic ash layers). The data is very robust and what results is a very detailed graph zig-zagging between warm and cold periods back in time. Not just one or two glacials and interglacials, but a myriad of them!

From analysing the Oxygen isotope ratios recorded in the foram tests, we can see many warm and cold stages throughout the past. (Image by Hannes Grobe Public Domain)
We know from the large fossils on land that there were hot and cold times. What the little foraminifera show us is there were more that we ever imaginged. These cold and warm times recorded by the ocean sediment correlates to the glacials and interglacials on land. The obvious question is what on Earth is creating such dramatic changes in the global climate?
The answer is, as always, not just one factor which contributes. And it is not just on Earth either. In the 1930s the Serbian geophysist and astronomer, Mulutin Milankovic, developed a theory that the Earth went through cycles of change as it travelled around the sun. Milankovic proposed that there were three main cycles. The first was Eccentricity which proposed a change in the Earth’s orbit around the sun being more elliptical in a 100,000 year cycle. Milankovic also proposed that the Axial Tilt varied in a 41,000 year cycle. The third cycle he called Precession where the ‘wobble’ of the Earth was quite extreme in a 21,000 year cycle. These three calculations are the Milankovic cycles. This theory based on calculations using observations in space has been backed up by the Oxygen Isotope record, correlating cycles to changes in the climate.
There are other factors too that can affect the climate. Sunspots on the surface of the sun can potentially reduce the amount of heat Earth recieves resulting in mini-ice ages. This is a relatively new theory and is still being debate whether or not sunspots have any effect. Something that can have quite dramatic effects on the climate is right here on our planet.
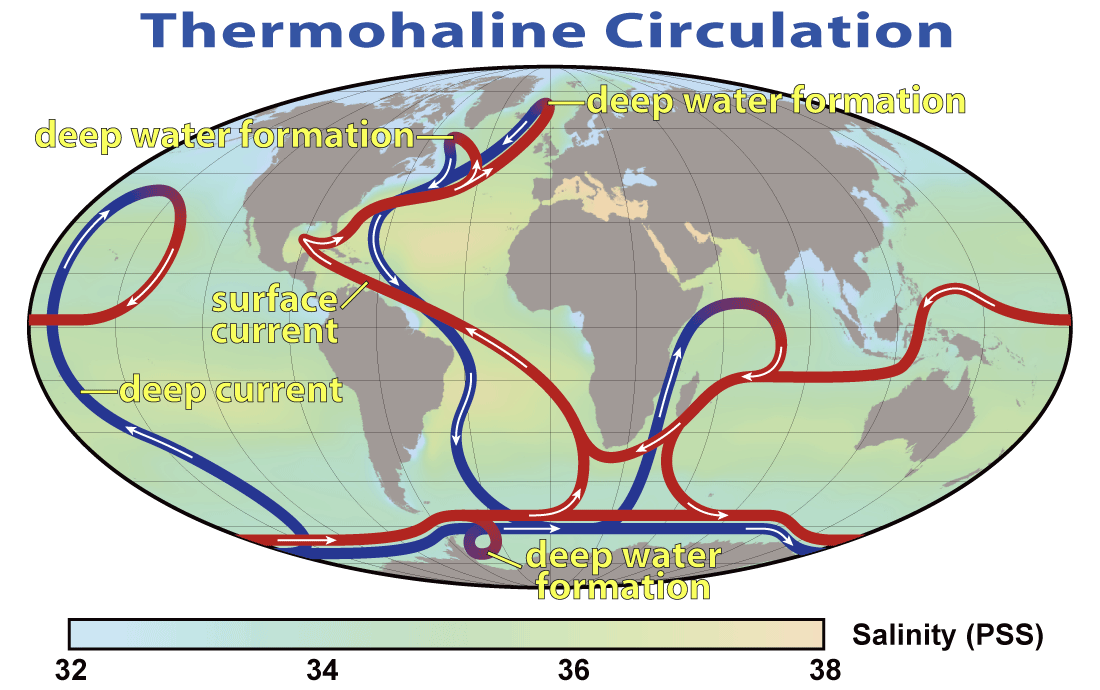
Unseen by us are warm and cold currents traveling around the planet in the oceans. The currents can change and if they do they can have dramatic effects on the climate. (Image by Robert Simmon, NASA. Public Domain)
Ocean currents travel the whole planet, with the warmer currents traveling nearer the surface, and the colder, dense currents sinking deep down. The warm ocean current from the Gulf of Mexico is what keeps Britain from literally freezing in the winter. These warm currents have not always been warm – melting of the ice caps can release large amounts of cold water into the oceans, disrupting the temperature of the ocean currents, and having a knock on effect on the climate.
Normally we end our Twilight Beast posts about the creatures extinction, or their current endangered status. Refreshingly, we don’t have to do that for this one. From the tiny shell of our little foraminifera, we have been able to look at the past in incredible detail. Hold a tiny foraminifera fossil in your hand and you will not notice the weight, or even feel it on your skin. What you will feel is finding yourself suddenly floating beneath the waves alongside this enigmatic little organism. You can see the weird ectoplasm unconsciously wave around, grabbing food particles. You zoom in even closer and watch as Oxygen atoms are absorbed into the test. Oxygen atoms that record the planets climate. This unassuming single celled organism is the key for time travel.
Written by Jan Freedman (@JanFreedman)
Further Reading:
Andrews, J T, (2000), ‘Dating Glacial Events and Correlation to Global Climate Change’, in Noller, Jay S. Et al. (eds), Quaternary geochronology: methods and applications, 2000, American Geophysical Union. [Book]
Benn, D. I. & Evans, D J A. (1998), Glaciers and Glaciation. Arnold Publishers. [Book]
Bond, G. et al. (1992), ‘Evidence for massive discharges of icebergs into teh North Atlantic Ocean during the last glacial period’, Nature. 60. pp.245-249. [Full article]
Bond, G, & Lotti, R. (1995), Iceberg discharges into the North Atlantic on millennial timescales during the last glaciation. Science. 267. pp.1005-1010. [Abstract only]
Curry, R. & C. Mauritzen, (2005). ‘Dilution of the northern North Atlantic in recent decades’, .Science 308: pp.1772–1774. [Abstract only]
Rabassa, J & Ponce, J F. 2013, ‘The Heinrich and Dansgaard-Oeschger climatic events during Marine Isotopic Stage 3: Searching for appropriate times for human colonisation of the Americas. Quaternary International. 299. pp.94-105. [Abstract only]
Rahmstorf, S (2003), ‘The concept of the thermohaline circulation’, Nature 421(6924): pp.699.
Rohling, E. J. et al.(2007). ‘High rates of sea-level rise during the last interglacial period’. Nature Geoscience 1: pp.38–42. [Abstract only]
Wright, J D. (2000), ‘Global Climate Change in Marine Stable Isotope Records’, in Noller, J. S. Et al. (eds), Quaternary geochronology: methods and applications, 2000, American Geophysical Union. [Book]




























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}