Head held down, the reindeer forced itself forwards. A strong, icy wind blasted against it’s thick hide. Straining to hear in the howling storm, there was no sign of the herd. Some moments earlier it had become separated from the group. Something had startled the reindeer. A wolf? Something else? The large four toes are spread out to allow this large deer to effortlessly move through the deepening snow, despite the oncoming stormy weather. The reindeer slowly presses onwards, although the top of the cliff is not sheltered and both the cliff and the reindeer take a battering from the blizzard. Down below, a small river cuts through an empty, white covered valley; all the animals are hibernating, or have migrated west. In the valley yellow lichen clings to the light grey hard rocks with a few mossy bushes poking out between the cracks. Visibility is reduced on the cliff top as the wind increases, but the reindeer blindly pushes onwards. The hole was covered by a thin blanket of snow. The reindeer couldn’t have known it was there. No animal could have. It snorted loudly as it fell, some thirty feet, down a dark shaft. From an awkward position, the reindeer shifted it’s head to watch a few snowflakes softly fall down from the small opening far above. It watched the last few clouds of breath dissipate into nothing, as eternal darkness slowly descended and the big reindeer closed it’s eyes one last time.
Around 15,125 years later (give or take 390 years), the skeleton of this unfortunate reindeer was found. The long shaft had ended in a small cavern. This breccia filled hole was excavated in 1964. The cavern was in Plymouth, Devon.

A beautiful reindeer (Rangifer tarandus) making it’s way across the mossy cold expanses in Sweeden. (Image from here)
A familiar mammal today, reindeer are one of the survivors of the Ice Age. Popularised by Santa’s original eight reindeer pulling his sleigh through the sky whereupon this one man delivered presents to every single house in the world in one night. The story of flying reindeer originates in Clement Moore’s 1823 poem ‘T’was the Night Before Christmas‘. Rudolf the ‘red nosed reindeer’ joins the group in the late 1930s. Clearly chosen for their strength and endurance during cold hard times, reindeer have become immortalised and symbolic as a part of Christmas. (I often wonder what people will think in 500 years time when they read stories about Father Christmas and his small, but trusty, herd of reindeer flying him across the world. Humans are quite good at making up stories.)
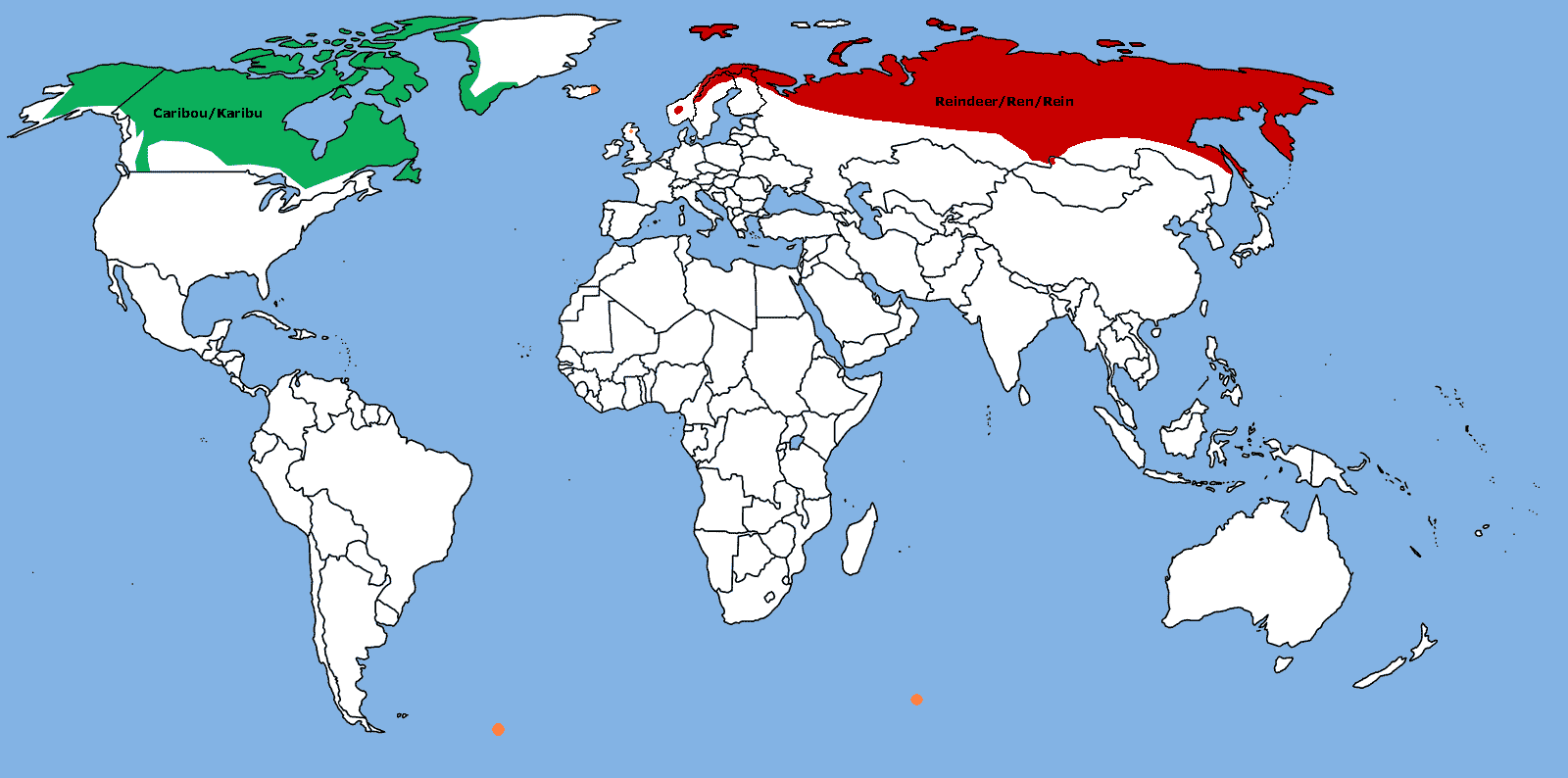
Today reindeer (Rangifer tarandus) are restricted to the Northern Hemisphere; Norway, Finland, Greenland, and Siberia, and to the east across Canada and Alaska. To the east, the herds of Rangifer tarandus are known as caribou, and across Eurasia, they are called reindeer. They are both the same species (they can mate and have fertile offspring). Reindeer may be slightly smaller than caribou which may be due to the more open landscape where the caribou live. These are herd animals, travelling together in huge numbers, which can be anything from a few dozen to a million strong. Migrating across flat snow covered plains to forested areas, reindeer are unique creatures with a very unique diet; they use their big, padded toes to dig in the snow to feed on lichens. Yes. Those flat rather non-descript algae/fungus symbiotic organisms. Lichens. Their strong curved incisors are perfect for nipping and scraping up this unusual food. Few other animals rely on lichen as a food; those that do include musk ox, Arctic hare and Arctic lemming. Although lichen is their main source of food in the winter seasons when other food is scarce, reindeer also eat moss, grass, and birch and willow leaves.

The modern day range of reindeer (and caribou). (Image from here)
So what was this reindeer doing making its way across Plymouth? Was it an isolated individual? Far, far from it. Reindeer were very abundant in Europe during the late Pleistocene.
Fossils of these majestic deer have been found in Germany dating back to the Early Middle Pleistocene (around 600,000 years ago), and shortly after fossils appear at sites in France. The climate was colder and drier during this period, allowing the range of the reindeer to be pushed much further south than it currently is. The climate during the entire Pleistocene was fluctuating like man-flu; there would be rapid periods of over the top hot times, followed by longer periods of real chilling coldness. During the colder phases, the reindeer travelled further south, following their habitat they favoured. In Britain the first evidence of them comes from Somerset’s Banwell Bone Cave, dating to around 83,000 years ago, along with some specimens found in Norfolk. After a small hiatus, remains of British reindeer can be found across many sites in the UK, from around 40,000 years ago to the end of the Twilight, 13,000 years ago. They spread so far south that fossils have been found in Kent’s Cavern, Devon, and in Plymouth.
Migrating across the cold Steppes of Europe along with other giants of the Pleistocene, reindeer were perfectly suited to these cold and tough environments. Mammoth, woolly rhinoceros, and horses enjoyed these colder Arctic times, along with much smaller creatures like the little Norway Lemmings (Lemmus lemmus), the tundra vole (Microtus gregalis) and the extinct water vole (Arvicola cantiana). The treeless landscape was dominated by lichens hanging on to the bare rocks, along with soft, bouncy light and dark green mosses, and low lying shrubs. Reindeer shared the flat expanse dotted by small dark forests with a surprising number of its relatives, including the magnificent giant deer (Megaloceros giganteus), red deer (Cervus elaphus), elk (Alces alces), roe deer (Capreolus capreolus) and fallow deer (Dama dama). With huge numbers of herbivores making their way across the landscapes, there was no shortage of predators. Wolves, cave lions, sabre tooth cats, leopards, and hyenas were all familiar carnivores in Europe until around 11,000 years ago.
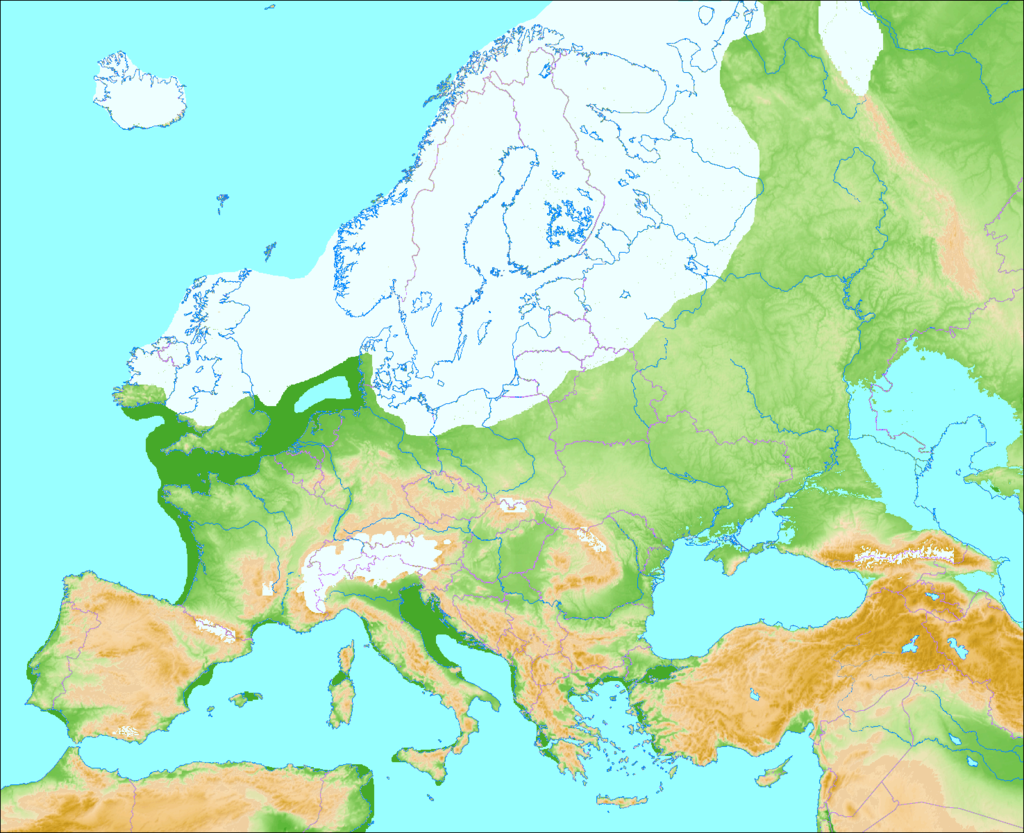
Something was happening that pushed reindeer away from Britain. At the height of the final glacial period in Britain, around 20,000 years ago (known as the Last Glacial Maximum), an enormous sheet of ice covered much of north western Europe, and most of Britain. This was a huge ice sheet and there was another one across the Atlantic covering much of North America, as far down to New York). With such huge quantities of water locked in the ice, the sea levels were much lower, and the English Channel was but a small meandering river surrounded by luscious grassy plains. Herds of animals moved with ease across non-existent geographical barriers. Hyenas, reindeer, Mammoths and other mammals from the Steppe moved freely across Spain, France, Italy and Britain. The ice, however, was not to last.

A nice illustration of the extent of the Ice Sheets during the last Glacial period of the Pleistocene. (Image from here)
Around 20,000 years ago, the ice began to melt. With a slowly warming climate, ice melted, sea level rose, and the vegetation across the landscape changed. The Mammoth Steppe that supported such a huge range of mega-fauna began to fade away. Many clung on to the disappearing Steppe environment, following north west as it slowly vanished. For those animals that stayed put, the environment they were suited to changed too much for them. Thick oak woodlands covered much of Britain, perfect for the smaller deer species, but not for reindeer. These wonderful animals became extinct in Britain around 13,000 years ago.
Their sheer numbers, and adaptation to extreme environments may have saved this Twilight Beast from extinction. Able to live in the coldest conditions, the few predators that picked off a few tasty reindeer made little impact to their overall numbers.
It appears humans fascination, and respect, for reindeer is more ancient than Father Christmas himself. Cave paintings at Cueva de las Monedas, in Spain include reindeer, demonstrating their presence in the area between 35,000 and 11,000 years ago. Capturing the essence of the animal on the wall inside a cave also shows that the artist saw something in this beast, something worthy of recording it on the walls. Fascinatingly, reindeer are one of the few species that do not appear on the famous painted caves at Lascaux, in France (even though reindeer bones have been found in the caves).
One of the most incredibly beautiful works of art made by human hands is a small piece of carved mammoth tusk found in Montastruc, France. Carved in unbelievable detail are two ‘crouching’ reindeer. The piece is called the ‘swimming reindeer’ as it looks as though they are swimming. This may not be a bad guess; Montastruc is a site which overlooks a river, so the carver may have witnessed reindeer swimming across. And reindeer, with their particularly large padded toes, are actually pretty good swimmers (although they would not swin streamlined like this, more of doggy paddle style). Another explanation could be that the shape of the mammoth tusk limited what the carver could actually carve. This one single sculpture, surviving for 13,000 years, gives us a glimpse into a long lost world, when reindeer were moving through France, and they captured the imagination of one carver.

Intricately carved from fragile mammoth tusk, these two reindeer look as though they are swimming, although this may be due to the lack of depth of a mammoth tusk. (image from the British Museum)
Written by Jan Freedman (@JanFreedman)
*Scroll past the references to read the full poem, T’was the night before Christmas introducing Santa’s reindeer for the first time by Clemet Moore (1823).
Further Reading:
Clark, P, U, et al. (2009). ‘The Last Glacial Maximum’, Science 325 (5941). pp. 710–4. [Abstract only]
Currant, A & Jacobi, R. (2001), ‘A formal mammalian biostratigraphy for the Late Pleistocene of Britain’, Quaternary Science Reviews. 20. 1707-1716. [Full article]
Jeffers, H. P. (2001), ‘Legends of Santa Claus’. Minneapolis: Lerner Publications. [Book]
Kurten, B. (1968), ‘Pleistocene Mammals of Europe’, The World Naturalist. [Book]
Montogomery, W, I, et al. (2014), ‘Origin of British and Irish mammals: disparate post-glacial colonisation of species introductions’, Quaternary Science Reviews. 98. pp.144-165. [Full article]
Stuart, A. J. (1977), ‘The Vertebrates of the Last Cold Stage in Britain and Ireland’, Philosophical Transactions of the Royal Society of London. 280 (972). pp.295-312. [Abstract only]
Stuart, A. J. (1982), ‘Pleistocene Vertebrates in the British Isles’, Longman. [Book]
Stuart, A. J. (1991). ‘Mammalian extinctions in the Late Pleistocene of Northern Eurasia and North America’, Biology Reviews. 66. pp.453-562. [Abstract only]
Sutcliffe, A. J. & Lewerne, B, (1977), ‘An unsolved mystery: the age of the almost destroyed human remains from Cattedown Cave, Plymouth, Devon’, Studies in Speleology. 3(1). pp.43-48.
Sutcliffe, A. J. (1985), ‘On the track of Ice Age Mammals’, British Museum (Natural History). [Book]
van Kolfschoten, T, et al. (2011), ‘A remarkable collection of Late Pleistocene reindeer (Rangifer tarandus) remains from Woerden (The Netherlands)’, Quaternary International. 238. pp.4-11. [Full article]
‘A visit from St Nicolas’
(also known as ‘T’was the night before Christmas’)
By Clement C Moore (1823)
‘Twas the night before Christmas, when all through the house
Not a creature was stirring, not even a mouse;
The stockings were hung by the chimney with care,
In hopes that St. Nicholas soon would be there;
The children were nestled all snug in their beds;
While visions of sugar-plums danced in their heads;
And mamma in her ‘kerchief, and I in my cap,
Had just settled our brains for a long winter’s nap,
When out on the lawn there arose such a clatter,
I sprang from my bed to see what was the matter.
Away to the window I flew like a flash,
Tore open the shutters and threw up the sash.
The moon on the breast of the new-fallen snow,
Gave a lustre of midday to objects below,
When what to my wondering eyes did appear,
But a miniature sleigh and eight tiny rein-deer,
With a little old driver so lively and quick,
I knew in a moment he must be St. Nick.
More rapid than eagles his coursers they came,
And he whistled, and shouted, and called them by name:
“Now, Dasher! now, Dancer! now Prancer and Vixen!
On, Comet! on, Cupid! on, Donner and Blitzen!
To the top of the porch! to the top of the wall!
Now dash away! dash away! dash away all!”
As leaves that before the wild hurricane fly,
When they meet with an obstacle, mount to the sky;
So up to the housetop the coursers they flew
With the sleigh full of toys, and St. Nicholas too—
And then, in a twinkling, I heard on the roof
The prancing and pawing of each little hoof.
As I drew in my head, and was turning around,
Down the chimney St. Nicholas came with a bound.
He was dressed all in fur, from his head to his foot,
And his clothes were all tarnished with ashes and soot;
A bundle of toys he had flung on his back,
And he looked like a pedler just opening his pack.
His eyes—how they twinkled! his dimples, how merry!
His cheeks were like roses, his nose like a cherry!
His droll little mouth was drawn up like a bow,
And the beard on his chin was as white as the snow;
The stump of a pipe he held tight in his teeth,
And the smoke, it encircled his head like a wreath;
He had a broad face and a little round belly
That shook when he laughed, like a bowl full of jelly.
He was chubby and plump, a right jolly old elf,
And I laughed when I saw him, in spite of myself;
A wink of his eye and a twist of his head
Soon gave me to know I had nothing to dread;
He spoke not a word, but went straight to his work,
And filled all the stockings; then turned with a jerk,
And laying his finger aside of his nose,
And giving a nod, up the chimney he rose;
He sprang to his sleigh, to his team gave a whistle,
And away they all flew like the down of a thistle.
But I heard him exclaim, ere he drove out of sight—
“Happy Christmas to all, and to all a good night!”
































{kind=link}
{kind=link}
{kind=link}
{kind=link}
.svg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}